|
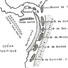
Revision of the historical collections of Pliocene-Pleistocene large mammals from Le Riège and Saint-Palais localities, near Pézenas (Southern France)
Published online: 23/06/2025
Keywords:
Hérault; Mammalia; Montpellier; Neogene; Quaternary
https://doi.org/10.18563/pv.48.1.e2
Abstract
Numerous “Quaternary” large-mammal fossils have been collected since the 1830s along the Le Riège stream, near Pézenas (Southern France). More than 120 specimens are stored in the collections of the Université de Montpellier (UM) under the name “Le Riège”. A major operation aiming at relocating the palaeontological collections of the University has made it possible to group together all the specimens of interest and launch their systematic revision for the first time. The fossils belong to the Reboul (1839; 51 samples) and de Christol (1865; 18 samples) Collections and 17 samples compose the Crochet & Ivorra Collection (1998). The remaining 38 samples have no mention about the exact time and location of their finding. We provide a critical inventory with literal transcription of inscriptions on specimens and historical labels. This revision confirms the presence of two distinct faunal assemblages under the name of “Le Riège”: Saint-Palais (Early Pliocene, MN14–15) and Le Riège sensu stricto (late Early Pleistocene, most likely MNQ19). The former assemblage, with coastal affinities, is composed of the ruminants Alephis sp. and Procapreolus cf. pyrenaicus, the rhinocerotid Pliorhinus megarhinus, the gomphotheriid Anancus arvernensis and marine mammals, all emblematic taxa for the Early Pliocene of Montpellier and Perpignan. The latter assemblage documents a late Early Pleistocene fluvio-volcanic sequence, yielding the bovid Bison (Eobison) spp., the cervid Eucladoceros cf. giulii, the hippopotamid Hippopotamus antiquus, the rhinocerotid Stephanorhinus etruscus, the equid Equus cf. altidens, and the elephantid Mammuthus cf. meridionalis, plus a few specimens of uncertain taxonomic affinities. This revision underscores the interest of revisiting historical collections and further provides a starting point for future research.
PV article infos
Published in 48-1 (2025)
|
PDF
S.I. Data
|
|
The Quaternary avifauna of Crete, Greece.
Published online: 01/09/1988
Keywords:
Avifauna; Crete; Quaternary; Systematics
https://doi.org/10.18563/pv.18.1.1-94
Abstract
Pleistocene bird fossils have been studied from nine localities on Crete. Part of this material was described earlier by the author (Weesie, 1982) and will not be treated here in extenso, the results will be incorporated. More than one third of the over 10,000 fossil bird bones available could be identified ; they were found to represent at least 65 bird species. The following species of the Pleistocene Cretan avifauna are new to the fauna of Crete : Branta ruficollis, Haliaeetus albicilla, Gyps melitensis, Aquila chrysaetos simurgh n. ssp., Ketupa zeylomensis, Aegolius funereus, Dendrocopos leucotos, Zoothera dauma, Turdus iliacus and Pyrrhula pyrrhula. The Pleistocene Cretan avifauna differs less from comparable mainland avifaunas than (fossil) avifaunas from oceanic islands do. Still, the Pleistocene Cretan avifauna has two qualities that are characteristic of island avifaunas : the almost complete absence of a group of birds (the Galliformes) and the presence of two endemic (sub)species : the giant eagle Aquila chrysaetos simurgh n. ssp. and the long-legged owl Athene cretensis (Weesie, 1982). The new subspecies is described in the present study.
These endemic birds of prey were found in association with their supposedly principal prey species (now extinct as well) : endemic mice for the owl and endemic deer for the eagle. Endemic mammals have been found in association with endemic birds of prey on many islands, not only in the Mediterranean. There is evidence that the size of endemic birds of prey becomes optimally adapted to their feeding on certain endemic mammals, especially rodents. Another characteristic of the Pleistocene Cretan avifauna is the great number of species of birds of prey. This appears to be a common characteristic of fossil avifaunas from caves on Mediterranean islands as well as from caves on the European mainland. However, we think that ecological conditions on Pleistocene Crete (especially the abundant presence of mice) helped to account for the high representation of birds of prey. Furthemore, the fossil avifauna enables us to draw some conclusions about the climate and vegetation on Pleistocene Crete : it is concluded that the climate was cooler than today and that Crete was largely covered with forests. Finally, the reasons for the extinction or disappearance from Crete of some bird species of the Pleistocene Cretan avifauna are discussed.
PV article infos
Published in Vol. 18, Fasc. 1 (1988)
|
PDF
|
|
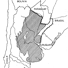
Mammals and stratigraphy of the continental mammal-bearing Quarternary of South America
Published online: 16/12/1984
Keywords:
Geochronology; Mammalia; Quaternary; South America; Stratigraphy
https://doi.org/10.18563/pv.14.ext
Abstract
Previous chronological arrangements of South American Quaternary land mammal faunas are appraised on the basis of current geological and paleontological data. Three South American late Pliocene-Pleistocene land mammal ages are conventionally recognized, from oldest to youngest, the Uquian, Ensenadan, and Lujanian ; all are defined on Argentine faunas.
The Uquian is based fundamentally and historically on the fauna from the Uquía Formation in Jujuy Province, northwestern Argentina. Important known formations in Argentina yielding Uquian Age faunas include the sub-surface Puelche Formation (or Puelchense) near the city of Buenos Aires, and the Barranca de Los Lobos and Vorohué Formations between Mar del Plata and Miramar, Buenos Aires Province. A tentative subdivision is propos-ed for the Uquian into three subages based on knowledge of the Mar del Plata-Miramar sequence, from oldest to youngest, the Barrancalobian, Vorohuean, and Sanandresian. In Argentina the Uquian is presently marked by the first known record of Scelidodon, Hydrochoeropsis, Ctenomys, Canidae, Ursidae, Gomphotheriidae, Equidae, Tapiridae, Camelidae, Cervidae, and the last known record of Thylatheridium, Thylophorops, Dankomys, Eumysops, Pithanotomys, Eucoelophorus, Hegetotheriidae, Sparassocynidae, and Microtragulidae.
The Ensenadan Age is based on the fauna from the Ensenada Formation near the city of Ensenada, Buenos Aires Province. In Argentina the Ensenadan is marked by the first known record of Lomaphorus, Neothoracophorus, Plaxhaplous, Cavia, Lyncodon, Lutra, Galera, Smilodon, Dicotyles, Lama, Vicugna, the last known record of Orthomyctera, and the only known record of Brachynasua.
Typícal beds of late Lujanian Age in Argentina consist of fluvial deposits occupying stream channels, and shallow basins, often incised into beds of early Lujanian (i.e. Bonaerian of early workers) and Ensenadan Age. The Lujanian Age is based on a fauna from beds along the Rio Luján, about 65 km west of the city of Buenos Aires, Buenos Aires Province. The Lujanian in Argentina is marked by the first record of Equus, Chlamyphorus, and Holochilus, and the last record of Megatherioidea, Glyptodontoidea, Arctodus (=Arctotherium), Smilodon, Litopterna, Notoungulata, Proboscidea, Equidae, Morenelaphus, and Palaeolama.
These land mammal ages are often difficult to recognize in other South American countries. The compositions of South American Pleistocene faunas vary with the environment. Some taxa were widely distributed in fossil deposits throughout the continent, but their occurrences need not reflect synchroneity. This is a result of changing climates and habitats in time. Consequently, proposed intracontinental correlations need confirmation based on magnetostratigraphy and a radioisotope time scale. Paleontologic characterizations of these land mammal ages (i.e. first and last record, and guide fossils) are useful for much of Argentina, but extensions to most of the other parts of South America are at best tenuous.
The majority of known non-Argentine Pleistocene faunas are believed to be Lujanian in age. Possible non Argentine early Pleistocene (Uquian) faunas include Ayo Ayo and Anzaldo in Bolivia, and Cocha Verde in southern Columbia. A possible middle Pleistocene (Ensenadan or early Lujanian) fauna is the Chichense of Ecuador. Paleomagnetic and radioisotopic date (MacFadden et al., 1983) clearly indicate that the greater part of the Tarija fauna (Bolivia) is Ensenadan in age.
The end of the Pleistocene and beginning of the Holocene in South America is marked by extinction of nearly all large mammalian herbivores and their specialized large predators. Radiocarbon age determinations suggest that large scale extinctions of megafauna occurred between 15,000 and 8,000 yrs. B.P. (years before present).
PV article infos
Published in Vol. 14, Ext (1984)
|
PDF
|